BioNB2220 CI Section
Effect of Conductance Changes on the
Hermissenda Photoreceptor
By Spencer Zhang
Introduction
One of my primary interest in neurobiology is how learning occurs in the nervous system. Therefore, I was interested in building a model of a learning system for my final project. To this end, I created a simulation of a photoreceptor in the mollusk Hermissenda crassicornis, which is directly associated with conditioned learning in the organism.
Classical conditioning
Hermissenda crassicornis provides an ideal neural system for studying how learning can occur in a brain. Although it has a very simple nervous system, it had been found to exhibit conditioned learning similar to that of more complex mammals. An untrained and naïve organism, when exposed to light, will attempt to move exhibit phototaxis, or move towards the light source. However, when exposure to light (the conditioned stimulus) is coupled with rotation (an unconditioned stimulus that discourages movement), the organism learns to associate the two stimuli with each other and is discouraged from further phototaxis. Experiments have shown that modifications to certain photoreceptors in the Hermissenda eye are sufficient to generate this conditioned behavior. How modifications to a photoreceptor can generate changes in behavior can be explained with the neural network below.
The neural network
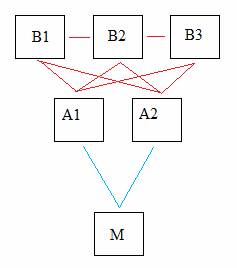
The Hermissenda eye is made up of two types of photoreceptors: Three type-B photoreceptors, which are the targets of the aforementioned modifications, and two type-A photoreceptors. The type-A photoreceptors stimulate a motor neuron responsible for the phototactic behavior in Hermissenda. In turn, the type-B photoreceptors form inhibitory synapses with the type-A receptors, as well as with each other. An observed result of classical conditioning is a broadening of the action potential produced in the type-B photoreceptor, which leads to a greater inhibition of type-A photoreceptors and consequently the motor neuron further down the pathway. Therefore, modifications to the type-B photoreceptor can generate a change in Hermissenda behavior in which phototactic behavior is discouraged.
The photoreceptor model

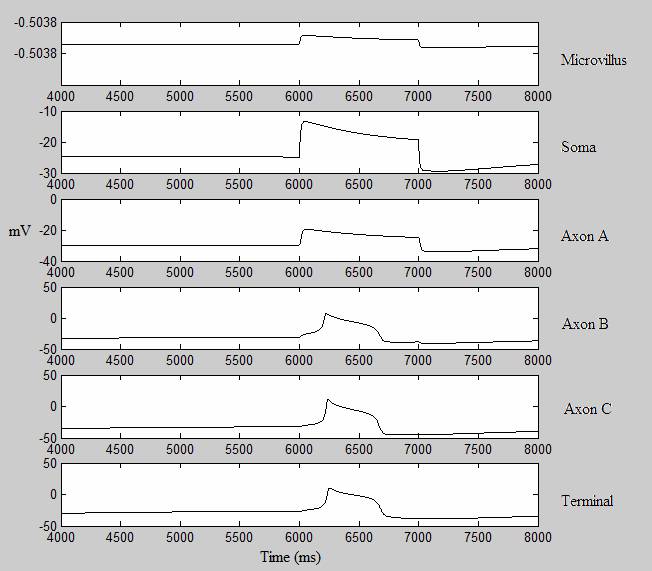
For this project, I created a multi-compartmental model of this type-B photoreceptor and focused on how changes in channel conductance affect the output at the synaptic terminal. The photoreceptor model is driven by the activities of seven ion channels (NA, H, KA, KV, CAT, CAS, and KCA), the leak current, as well as the current between the compartments themselves. The channels are represented by Hodgkin-Huxley formulations. The steady state and time constants for the activation and deactivation functions of each channel were taken from Flynn’s paper. At the moment, the model is set up to study the effects of changes in channel conductance, so the main inputs taken by hermis.m are the percentages of each channel’s conductance. For example, hermis([1,1,1,1,1,1,1]) produces the spiking expected in a naïve organism. The code hermis([1, 1, 0.5, 0.5, 1, 1, 1]) runs a trial in which the conductances of the potassium channels, KA and KV, are reduced to 50% of their original conductances.
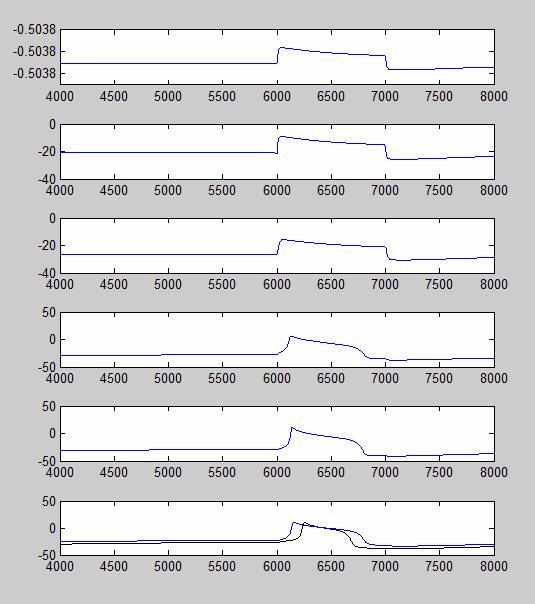
In the test shown below, +50 uA of charge was injected into the soma of the photoreceptor, creating a generator potential. The potential is then carried to the axons, where the generator potential is converted to an action potential and finally carried to the synaptic terminal. The difference between the voltage patterns in the six departments is due to the difference in what ion channels are present in each compartment. For example, the conversion of generator potential to action potential can only occur in axons B and C due to the fast sodium channel, which is only present in those two compartments. The simulations are actually started at t=0, but only the voltage after t=4000 are recorded here. I gave the model a 6000 msec delay between the start of the simulation and the current injection to let it reach a steady state before spiking. Under some conditions, the axons will spontaneously produce an action potential at the start of the simulation, and without a sufficient delay the hyperpolarization period of the first action potential will prevent the second spike from firing.
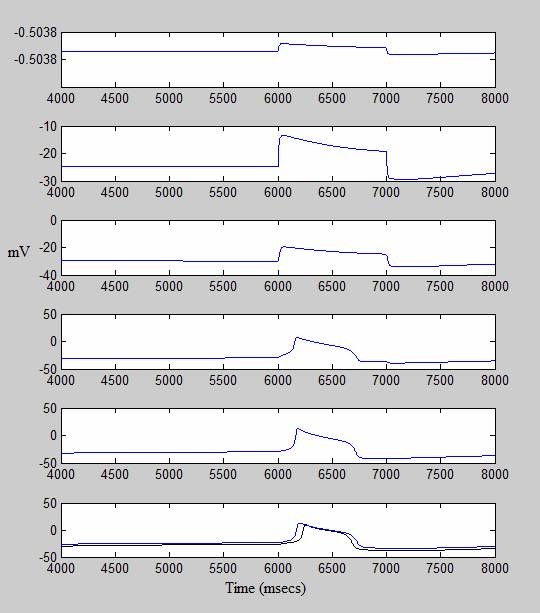
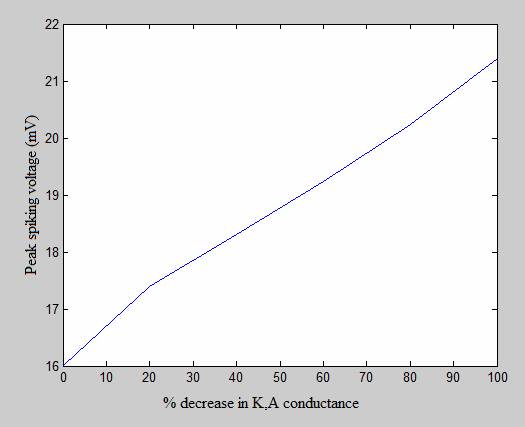
The following graphs show an experiment in which I decreased the conductance of the Type-A potassium channel (K,A), the relatively fast K+ channel. The graph on the bottom left compares the action potential produced under normal conditions (drawn in black) with the action potential produced when the conductance of the K,A channel is reduced to zero. The graph on the right shows how the peak spiking voltage (in mV) increases as K,A conductance decreases. (The values on the X-axis refer to the percentage by which the K,A conductance is decreased. Therefore, 0 refers to a trial under normal conditions and 100 refers to a trial where the channel is completely disabled.
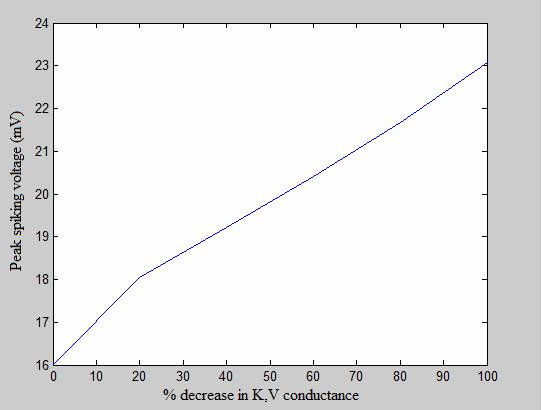
The graphs below show a similar experiment conducted on the delayed rectifier (K,V) channel, with similar results. These experiments show that as the conductance of potassium channels decreases, the peak voltage of the action potential increases, the spikes reaches its peak earlier and lasts for a longer period of time, all of which is consistent with empirical findings. The graphs below show that K,V has an even spike-broadening effect than the K,A channel.
Conclusion
From the data produced from the simulation, I can conclude that the simulation accurately portrays the response of the Hermissenda photoreceptor to decreases in the conductances of potassium channels. There is still a number of characteristics that needs to be verified, in particular that of the calcium-dependent potassium channel (KCA). While the channel itself has been implemented, changes to the channel conductance do not produce the same effect as have been produced in Flynn’s paper. In future studies, I would like to observe how Hermissenda’s visual network responds as a whole to modifications to the type-B photoreceptor, as well as the role of lateral inhibition between the three type-B photoreceptors. It is probable that learning in more complex mammalian organisms employ a much more complex mechanism than that used by Hermissenda. However, study of this simple learning system has given us a clear picture of how a number of simple modifications to a neuron can produce the effects of conditioned learning.
References
Flynn, M. Cai, Y. Baxter, DA. Crow, T. “A Computational Study of the Role of Spike Broadening in Synaptic Facilitation of Hermissenda” (2003). J. Comp. Neuro. 15: 29-41.
Crow, T. “Cellular and Molecular Analysis of Associative Learning and Memory in Hermissenda” (1988) TINS. 11:4:136-142.